
Immunoglobuliner forkortes IgX hvor X angiver typen. Det kaldes også for antistoffer. Antistoffer føres med modermælken ved amning og således er amning med til at styrke immunforsvaret.
Den store diversitet af antistoffer i vores repertoire skyldes følgende faktorer:
- Mange forskellige V, D, og J gener (Germline diversitet)
- Tilfældig rekombination af V-, J-, og D-regionerne (Kombinations diversitet)
- ”unøjagtig” aflæsning af D/J området (Junktional diversitet)
- tilfældig kombination af tung og let kæde
- Somatisk hypermutation
Man skelner mellem monoklonale antistoffer og polyklonale antistoffer. Monoklonale antistoffer har samme batch reaktivitet og kommer fra mus, kanin og mennesker, mens polyklonale antistoffer har forskellig batch reaktivitet og kommer fra kanin, ged, hamster, mus, kylling og andre.
Et antistofmolekyle kan betragtes som en B-celle receptor, der er secerneret fra B-cellens effektorcelle, plasmacellen. Antistoffet fungerer som to molekyler i eet. Dette gælder ikke blot for den strukturelle opbygning, men også funktionelt. Et antistof har en genkendelses- og bindingsevne i forhold til sit antigen, en egenskab der strukturelt er beliggende i Fab-delen.
Struktur
Antistofferne benævnes også immunglobuliner (Ig). Elektroforetisk tilhører de den fraktion af serumproteinerne, som benævnes γ-globuliner. Uafhængigt af antistofspecificitet kan immunglobulinerne inddeles i fem forskellige klasser: IgM, IgD, IgG, IgE og IgA. Denne inddeling beror på strukturelle forskelle i den konstante del af immunglobulinernes tunge kæder. Benyttes immunglobulin som antigen ved injektion til en anden art (hvis der for eksempel indsprøjtes humant immunglobulin i en kanin) vil der produceres antistoffer med specifik reaktionsevne mod hver enkelt immunglobulinklasse. De fem immunglobulinklasser findes hos alle individer og benævnes derfor isotyper.
Immunglobulinklassernes molekylstruktur er iøvrigt meget ensartet. Det drejer sig om glykoproteiner med en 4-kædet grundstruktur, bestående af to lette kæder (L = light, MW 25 kD), og to tunge kæder (H = heavy, MW 55 kD).
De enkelte kæder er knyttet sammen af disulfidbindinger og af ikke-kovalente kræfter. Bindingsområdet for antigen (paratopen) er lokaliseret til den N-terminale del af molekylet og medinddrager både den lette og den tunge kæde. Hvert antistofmolekyle har derfor to identiske bindingssteder for antigen – dog har et IgM-molekyle 10 bindingssteder og et IgA-molekyle optræder ofte som en dimer med 4 bindingssteder. Aminosyresekvensundersøgelser af forskellige humane myelom-proteiner har vist, at de første ca. 110 aminosyrer regnet fra den N-terminale ende udviser en ret udtalt variabilitet. Derfor benævnes de N-terminale dele af H kæden og L kæden de variable dele, forkortet henholdsvis VH og VL. Den tunge kæde har herudover tre (IgG, IgA, IgD) eller fire (IgM, IgE) konstante områder (CH1, CH2, CH3, CH4) Den lette kæde omfatter eet konstant område, CL. Det typiske antistofmolekyle har inter-kæde disulfidbroer som sammen med ikke-kovalente kræfter holder kæderne sammen. Molekylets opbygning og funktioner fremgår blandt andet af undersøgelser med enzymerne pepsin og papain, som spalter Ig-molekylerne på karakteristisk måde. Papain spalter molekylet i tre fragmenter af omtrent samme størrelse: to identiske Fab-fragmenter (affragment antigen-binding) og et Fc-fragment (affragment crystallizable). Ved pepsinspaltning dannes et dimert Fab.
Inddeling
Immunoglobulinerne kan inddeles i de fem subtyper:
- IgM
- IgD
- IgE
- IgA
- IgG
De forskellige isotyper for tunge og lette kæder er til stede hos alle individer inden for en art. Der eksisterer imidlertid yderligere for de konstante dele af immunglobulinkæderne områder, hvor der inden for samme art (for eksempel hos mennesket) er alternative aminosyresekvens-muligheder. Sådanne alternative former nedarves Mendelsk arvegang|Mendelsk og kaldes allotyper. For menneskets IgG kendes ca. 25 allotyper (Gm typer), for IgA tilsvarende to Am-allotyper. Hvad L-kæderne angår kendes tre allotyper for kappa-kæden, såkaldte Inv-allotyper. Der er ikke fundet allotyper for lambda-kæden.
Idiotyper kan defineres som de strukturer i den variable del af den lette og den tunge kæde, som betinger det enkelte antistofs specificitet for et bestemt antigen (paratopen). Det er muligt inden for en dyreart at fremstille anti-idiotypiske antistoffer ved at immunisere med et specifikt antistof. Herved kan dannes antistoffer mod den antigene del af paratopen (idiotopen). Fab delen af et anti-idiotypisk antistof må have en vis lighed med det antigen, det oprindelige antistof er rettet imod. I mange tilfælde vil anti-idiotypiske antistoffer være blokerende (hæmmende) for bindingen mellem antigen og antistof. Forekomsten af et netværk af idiotyper og anti-idiotyper inden for immunglobulinerne kunne være en fundamental egenskab ved B-lymfocytsystemet. Denne ide er fremsat af Niels Jerne, men det har dog været vanskelig at demonstrere betydningen af dette.
Den grundlæggende strukturelle enhed i immunglobulinmolekylerne er det såkaldte immunglobulindomæne. Immunglobulindomænet omfatter en sekvens på ca. 110 aminosyrer og har en molekylvægt på omkring 12,5 kD. Det er en kompakt, globulær struktur med en såkaldt ß-struktur, opbygget som antiparallelle løb af aminosyresekvenser i to planer. De to planer er holdt sammen af en disulfidbinding.
Den tunge kæde af et IgG molekyle er sammensat af fire sådanne domæner (VH og CH1-3) og den lette kæde af to domæner (VL og CL). Evolutionsmæssigt forestiller man sig, at de enkelte immunglobulin-domæner er opstået ved genduplikation ud fra et primitivt domæne-gen. Det er i denne forbindelse interessant, at andre af immunsystemets molekyler er opbygget af tilsvarende domæner: fx MHC Klasse I og Klasse II molekylerne og ß-2 mikroglobulin, samt den antigen- specifikke T-celle receptor.
En såkaldt J-kæde syntetiseres af den antistofsecernerende celle. Den er knyttet til den konstante del af α- eller µ-kæderne og indgår i polymerisationen af IgM og IgA og bindingen af secretory component (SC). J-kæden har en molekylvægt på 15 kD. Strukturelt har den ingen relation til immunglobulinets domænestruktur.
Ud over J-kæden kan der til IgA og til IgM være knyttet en anden polypeptidkæde, nemlig den såkaldte Secretory Component (SC). SC har en molekylvægt på 60 kD og syntetiseres ikke af de antistofproducerende celler, men af epitelceller. Efter syntesen bindes dimert IgA til en Fc-receptor (polyIg receptoren) på den basale side af slimhindernes epitelceller, hvorefter IgA transporteres ind i epitelcellen. Inde i epitelcellen bliver en del af Fc-receptoren klippet af, mens en del, SC, resterer, bundet til IgA’s J-kæde. SC har således betydning for en selektiv passage af IgA gennem epitelcellen til de eksterne sekreter hvorefter komplekset af IgA og SC bliver afgivet til sekreterne. I slimhindesekretet virker SC som en hæmmer af proteolytisk nedbrydning af immunglobulinet. IgM kan på samme måde som IgA transporteres ud i lumen via polyIg receptoren.
Antistoffernes isotyper defineres af de konstante områder af de tunge kæder. Hver isotype kodes for af et separat konstant-område gen. Disse gener ligger i et cluster 3’ for de variable gener. Når et variabelt område er produktivt rearrangeret udtrykkes det først med µ og ∂ tunge kæder, men det samme variable område kan senere udtrykkes sammen med en hvilken som helst af de andre isotyper. Denne proces kaldes isotype switch, og består i at DNA rearrangeres, så det variable område placeres 5’ i forhold til de forskellige konstant-område gener. Isotype switch foregår kun i antigen-stimulerede B-celler, kræver T-celle hjælp og kan altså bevirke at det samme antistof (den samme specificitet) kan have eller få forskellige funktionelle egenskaber via udskiftning af Fc (knyttet til de forskellige isotyper).
IgM
IgM molekylet er opbygget af fem identiske grundenheder, som er koblet sammen via S-S-bindinger og forsynet med J-kæde. Molekylvægten af IgM er ca. 900 kD, og et enkelt IgM molekyle har 10 antigen bindingssteder. Funktionelt virker IgM dog nærmest som en pentamer, sandsynligvis af steriske grunde. IgM antistoffer har på grund af de mange valenser en stærk evne til at virke agglutinerende såvel over for bakterier og virus som over for erythrocytter, og IgM antistoffer kan påvises meget tidligt i forløbet af en immunisering (infektion). IgM antistoffer er komplementbindende, og de betragtes som en væsentlig barriere i det tidlige forløb af invasion af mikroorganismer. Generelt er affiniteten af det enkelte bindingssted lav, men muligheden for multivalent binding medfører en høj funktionel affinitet (aviditet).
IgD
IgD indholdet i serum er meget lavt, det nedbrydes let af proteaser, og serum IgD synes ikke at have nogen speciel funktion. Membranbundet IgD ses ofte sammen med IgM og/eller IgG, og IgD synes at spille en rolle ved den ontogenetiske udvikling af B-lymfocytten (den naive B-lymfocyt udtrykker både IgM og IgD).
IgG
IgG er kvantitativt det helt dominerende immunglobulin i serum. Det er i mange henseender den vigtigste klasse af immunglobuliner. Antistoffer af IgG-klassen ses som nævnt typisk efter flere udsættelser for antigen (det sekundære immunsvar). De forskellige IgG-subklasser har forskellige egenskaber (bl.a. m.h.t. evnen til at aktivere komplementsystemet).
IgA
IgA findes i to former, en monomer form i serum og en dimer form i sekreter. IgA i sekreter synes at spille en væsentlig rolle for beskyttelse mod indtrængen af mikroorganismer via slimhinder. Afgivelse af IgA til slimhindeoverfladen er betinget af forekomsten af J-kæde og SC. Specifikke IgA antistoffer findes ofte i højere koncentration i sekreter end i serum, fordi syntesen af IgA foregår lokalt. IgA antistoffer aktiverer ikke den klassiske komplementaktiveringsvej, men kan virke neutraliserende på toksiner eller forhindre indtrængen af mikroorganismer ved blokering af adhæsionsstrukturer på mikroorganismerne.
IgE
IgE findes hos normale i forhold til de andre klasser af immunglobuliner i meget lav koncentration (se skema). Koncentrationen af IgE stiger ved visse parasitære sygdomme, specielt ved invasion af orme, ved visse immundefekter og ved nogle kroniske, IgE-betingede allergiske tilstande. IgE antistoffer har evnen til at bindes til receptorer (specielt FcεRI) på mastceller og eosinofile og basofile granulocytter via IgE’s Fc-region. De er af stor betydning ved type I allergiske sygdomme.
Antistoffernes diversitet
Den samlede diversitet af antistoffer er anslået til i størrelsesorden 1012 forskellige antistofmolekyler – et utroligt stort antal taget i betragtning det begrænsede genetiske materiale, der anvendes. Diversiteten skyldes flere mekanismer:
- I det enkelte individs genom findes talrige gen-segmenter, svarende til variable områder (V, (D), J), og er således en arvelig kilde til diversitet
- Diversiteten er nu et resultat af en dannelse af et komplet genområde svarende til det variable område ved tilfældig kombination af V, D, og J-segmenter.
- Variabilitet ved sammenføjninger mellem segmenter øges ved upræcis sammenføjning og ved indsættelse af såkaldte P- og N-nukleotider.
- Let og tung kæde sammenføjes ved en tilfældig proces og giver yderligere diversitet
- Endelig bevirker det senere fænomen, somatisk hypermutation, en yderligere udvidelse af diversiteten. Somatisk hypermutation er T-celleafhængig og foregår i kimcentre i perifere lymfoide organer.
Den bemærkelsesværdige diversitet blandt antistof molekylerne er konsekvensen af en højt specialiseret mekanisme, ved hjælp af hvilken de immunglobulin-gener der udtrykkes i en given celle er sammensat ved hjælp af DNA re-arrangeringer som sammensætter to (eller tre) gensegmenter hvorved der dannes en variabel-region under B-cellens udvikling. Senere switch-mekanismer kan sætte denne variable region sammen med enhver af generne for de konstante områder og på denne måde udvikle antistoffer hørende til hver af de fem isotyper. Disse processer foregår i knoglemarven under modningen, og når en naiv B-celle forlader knoglemarven udtrykker den altså én bestemt specificitet.
For den tunge kædes vedkommende er der yderligere mellem V-regionen og J-regionen indskudt en D-region.
Denne proces indebærer at et Ig-gen ikke oprindelig er ét langt DNA-stykke, men at det sammensættes af mindre segmenter. Det er naturligvis den variable del af molekylet der her er interessant, altså den del der koder for det første domæne på ca 110 aminosyrer: Gensegmenterne er for den tunge kædes vedkommende fordelt på tre områder, et V-område, et D-område og et J- område, og for den lette kædes vedkommende af to områder, et V-område og et J-område. Men der er mange individuelle segmenter, for den tunge kæde er der af V-områder ca 50, der er ca 25 D-områder, og der er 6 J områder.
Re-arrangeringen (her af den tunge kæde) består så i at først eet (tilfældigt) D-område ud af de 25 bliver sat sammen med eet (tilfældigt) J-område (af de 6) – og der er jo så 150 muligheder. Og hver af de 150 muligheder kan så knyttes sammen med eet (tilfældigt) af de 50 V-områder. Det giver i alt ca 7.500 muligheder.
For den lette kæde bliver der efter samme princip ca. 350 forskellige muligheder. Når så til sidst een tilfældig tung kæde knyttes sammen med een tilfældig let kæde bliver der i alt ca 2.6×106 forskellige immunglobuliner. For enderne af de forskellige gensegmenter er en såkaldt rekombinations signal sekvens (RSS), bestående af en heptamer-23 eller 12 basepar-en nonamer.
Funktioner
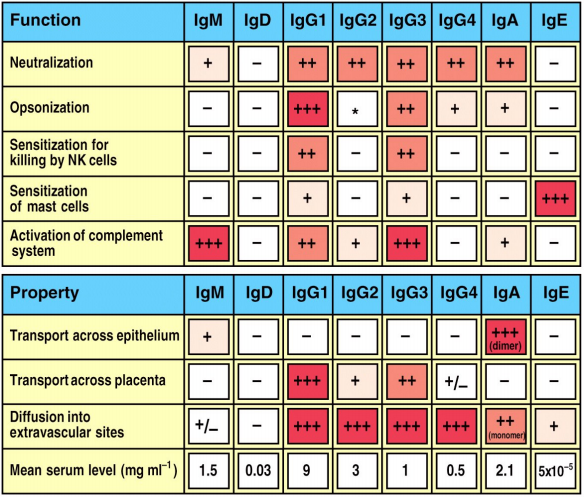
De forskellige antistof klasser og subklasser har forskellige egenskaber og funktioner – se billede.
Antistoffer kan hæmme virus og bakterie infektion (neutralisering). Ved virus binder antistoffet til virussen og forhindrer den i at binde til virusreceptorer på celleoverfladen. Ligeledes binder antistoffer til bakterier og forhindrer kolonisering og uptake i cellen.
Herudover kan antistoffer også hæmme mikroorganismers spredning og blokere for toxiner. Spredningen stoppes netop ved at antistoffer binder til mikroorganismer og forhindrer dem i at komme ind i andre celler. De kan også binde til toxiner og forhindre dem i at binde til deres receptorer.
Kun i visse tilfælde er et antistofs bindingsevne den væsentligste effektormekanisme ved et antistofs funktion, men et eksempel på dette er antistoffers neutralisation af et toksin som tetanustoksin eller difteritoksin – her er bindingen af antistof til antigenet i sig selv beskyttende for en organisme eller et antistofs binding til en struktur på en mikroorganisme, der er nødvendig for mikroorganismens interaktion med en targetcelle.
Kapselbærende bakterier
Nogle bakterier har kapsler der beskytter dem mod fagocytose, idet de ikke kan optages af neutrofile granulocytter. Binding af antistof (IgG) aktiverer komplementsystemet og binding af komplementfaktor C3b til bakterien. Nu kan bakterien optages ved binding til Fc-receptorer og komplementreceptorer. Nu kan granula fusionere med fagosomet og frigive toksiske oxygenholdige stoffer der dræber bakterien.
Drab af mikroorganismer
Antistof-coatede mikroorganismer kan binde til Fc-receptorer på fagocytter. Dette signalerer fagocytose af hele cellen og drab inde i cellen.
Også Natural Killer Cells (NK-celler) kan dræbe antistof-coatede mikroorganismer.
Inflammation
Krydsbinding af IgE på mastcelleoverfladen medfører frigivelsen af inflammatoriske mediatorer, såsom histamin.
Mekanisme
For den beskyttende funktion af antistoffer over for patogene mikroorganismer er det i mange tilfælde en forudsætning, at bindingen til den patogene mikroorganisme udløser en række effektorfunktioner, som alle foregår via antistoffets hale-del (Fc-delen). Disse effektorfunktioner udløses via komplementaktivering og binding til Fc-reptorer på celleoverflader (fagocyterende celler, NK-celler, basofile granulocytter og mastceller). Endelig er antistoffernes evne til at passere visse biologiske membraner en funktion af deres Fc-del, for eksempel fordøjelseskanalens og respirationsvejenes slimhinder, og overgangen af immunglobuliner til fosteret.
Immunglobuliner eksisterer både som overflade-receptorer for antigen på B-celler og som secernerede serum proteiner. Den overfladebundne (transmembran-form) og den secernerede form af et immunglobulin fra een bestemt B-celle har samme antistofspecificitet, og forskellene mellem dem beror på forskellig tung-kæde afslutning af transkription og påfølgende RNA processering. Dette er et væsentligt punkt i klon selektions teorien. Udtryk af immunglobulin i cellemembranen og signal-transduktion efter binding af antigen kræver at den tunge kæde er tilknyttet to andre membran-proteiner, Igα og Igβ. Disse to molekyler danner et kompleks i cellemembranen. Krydsbinding af dette kompleks er nødvendigt for at aktivere de intracellulære enzymer, som initierer den række af biokemiske processer, der fører til aktivering af protein kinaser og til en øgning af intracellulært calcium. Antigener som ikke i sig selv kan krydsbinde overflade immunglobulin eller kan stimulere via en mitogen effekt (TI-antigener) kan ikke aktivere B-celler uden hjælp fra T-celler. Aktiverede komplement komponenter udgør et specielt signal, og virker ved at bindes til komplement-receptorer (CR2, også benævnt CD21) på B-cellen, som danner et kompleks med CD19 og CD81 i cellemembranen og virker forstærkende på signaltransduktionen fra BCR.
Vi kan lave antistoffer mod stort set alle molekylære strukturer. Begrænsningen er tolerans – men den kan i de fleste tilfælde brydes. En anden begrænsning er, at det antistofferne skal reagere med skal være tilgængelig, skal kunne “ses”.
Et individ kan lave i størrelsesorden 1012 forskellige antistoffer, og dette bygger på et system af gensegmenter, som kan kombineres på mange måder – men også andre mekanismer bidrager til hvad vi kalder antistof repertoiret. Et væsentligt element i udnyttelsen af dette store repertoire er fænomenet recirkulation, som bevirker at de meget få lymfocytter der er specifikke for et bestemt antigen kan komme i kontakt med antigenet (i en lymfeknude, i milten, eller i MALT-en).
De modne, men naive lymfocytter (har aldrig set ”deres” antigen) kommer fra thymus (T-lymfocytter) og fra knoglemarven (B-lymfocytter). Langt de fleste møder aldrig ”deres” antigen – og dør – men der dannes til stadighed nye naive lymfocytter.
En epitop (begreb indført af Niels Jerne) er den struktur på antigenet som indgår i interaktionen med antistoffet. Når et antigen denatureres ændrer dette antigenisiteten, som kan defineres som en egenskab ved antigenet, det antistoffer kan være rettet imod. Begrebet immunogenisitet siger noget om hvor let det er at lave antistofferne i et bestemt individ. Der er her bl.a. genetiske faktorer, der spiller ind, men også den kemiske struktur af antigenet spiller en stor rolle.
Når et antigen kommer ind i kroppen, skal det for at udvikle et immunsvar først transporteres til et lymfatisk organ – og i denne proces er der et sammenspil mellem de tre celletyper, der er nødvendige for et antistof-svar: den specifikke B-lymfocyt (som når den er modnet bliver til en plasmacelle, der producerer antistof), den specifikke T-lymfocyt (som skal være hjælpecelle for at B-lymfocytten kan blive stimuleret, og endelig den antigenpræsenterende celle (dendritcelle, makrofag), som dels deltager i transporten af antigenet til det lymfatiske organ, og dels præsenterer antigenet for T-lymfocyt.
Fremstilling
Forskellige dyrearter har hver deres fordele og ulemper når de anvendes til antistoffremstilling:
- Kaniner – er gode til fremstilling af antistoffer til immun-præcipitationsteknikker
- Mus – er gode til immuniseringsforsøg – og til fremstilling af monoklonale antistoffer
- Større dyr – er gode til fremstilling af store mængder antiserum
- Høns – er gode p.gr. af den store udviklingsmæssige afstand fra pattedyr.
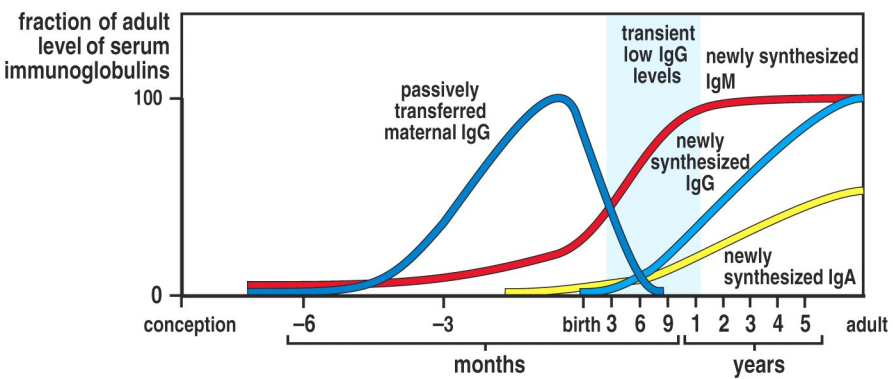
Neonatal immunforsvar
Spædbørn fødes med et antistoffer fra moderen, idet antistoffer kan passere placentamembranen. Ved fødslen danner spædbarnet stort set ikke selv antistoffer. I slutningen af første måned, er andelen af antistoffer fra moderen halveret. Herefter begynder babyen selv at danne antistoffer, og når barnet er mellem 12 og 20 måneder er antistofniveauet normalt.
Selvom antistofniveauet falder kort efter fødslen, beskytter moderens antistoffer mod de fleste børnesygdomme de første 6 måneder, herunder mæslinger, difteri og polio. Derfor er det oftests ikke nødvendigt at vaccinere mod disse sygdomme før denne alder. Nogle sygdomme som kighoste er barnet dog ikke beskyttet imod, og derfor er vaccination nødvendigt tidligt.
Sidst opdateret 19. maj 2023